
Noah Benson, Ph. D.
Depts. of Psychology & Neurology
University of Pennsylvania
![]() Research
Research
My research is focused on examining and modeling the neural correlates of vision in the retina and the visual cortex. I am interested abstractly in the question of how perception arises from the reduction and reorganization of the many millions of bits of information that are observed by the retina as this information passes through the visual pathway. Currently, I am studying both the organization of color in the retina and early visual pathway as well as the retinotopic organization of striate and extrastriate cortex. Download my CV.
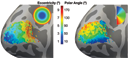 |
Retinotopic Organization of V1, V2, and V3 Area V1, the primary visual cortex, is the earliest cortical visual area. Our previous research (Benson |
|
|
Benson NC, Butt OH, Brainard DH, Aguirre GK (2014) Correction of distortion in flattened representations of the cortical surface allows prediction of V1-V3 functional organization from anatomy. PLoS Comput. Biol. Submitted. Benson NC, Butt OH, Datta R, Radoeva PD, Brainard DH, Aguirre GK (2012) The retinotopic organization of striate cortex is well predicted by surface topology. Curr. Biol. 22(21): 2081-5
|
||
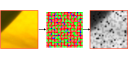
Learning Cone Types in the Retinal Mosaic In order for color vision to function, the brain must have some information about the types (L, M, or S) of the cones that are firing in response to any given scene. There is, however, no known biochemical mechanism by which the brain can learn the type of L and M photoreceptors. We ask whether the information inherent in the firing of photoreceptors in response to natural scenes is sufficient to type the retinal mosaic using an unsupervised learning algorithm. |
 |
|
Benson NC, Manning JR, Brainard DH (2014) Unsupervised learning of cone spectral classes from natural images. Submitted. Benson NC, Brainard DH (2012) An unsupervised learning technique for typing cones in the retinal mosaic. Talk presented at the 12th Annual Vision Sciences Society (VSS) Meeting. 11-16 May.
|
||
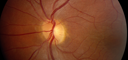 |
Drusen Sensitivity in Macular Degeneration The research on the sensitivity of hard drusen in early age-related macular degeneration (AMD) is split by evidence that they both are and are not less sensitive to stimuli displayed over their portion of the retina. We are working to shed light on this by analyzing both the sensitivity of drusen in AMD patients and genetic factors which may contribute to their relative sensitivity. |
|
|
|
||
My graduate research focused on explaining and characterizing the dynamics of proteins. I analyzed proteins in our Dynameomics database to examine dynamics of all proteins broadly using wavelet analysis, flexibility analysis, and graph theory. I additionally examined a smaller number of methyltransferase proteins with single nucleotide polymorphisms (SNPs) to examine how mutations affect protein dynamics.
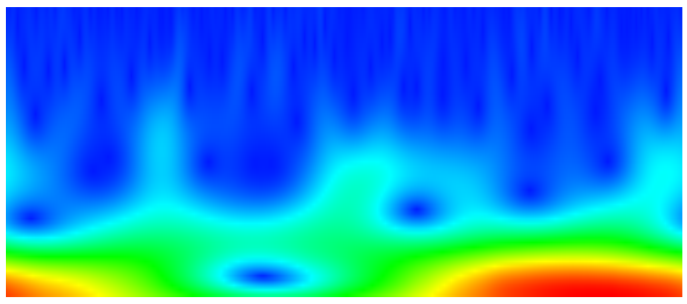 |
Wavelet Analysis of Protein Motion The motion of proteins is often difficult to interpret, and events of interest in a protein simulation can be difficult to locate. We have used wavelet analysis as a means of quickly scanning simulations for rearrangements and subtle changes in a protein that have physical significance. |
|
|
Benson NC, Daggett V (2012) Wavelet analysis of protein motion. Int. J. Wavelets Multi. Inf. Process. 10: 1250040-59. Benson NC, Daggett V (2012) A comparison of methods for the analysis of molecular dynamics simulations. J. Phys. Chem. B 116(29): 8722-31.
|
||
Chemical Environments: Protein Graphs Much research has already found applications of rudimentary graph representations of proteins. By representing similar chemical regions of a protein as nodes in a graph, I hope to simplify the observation of chemical changes over time. |
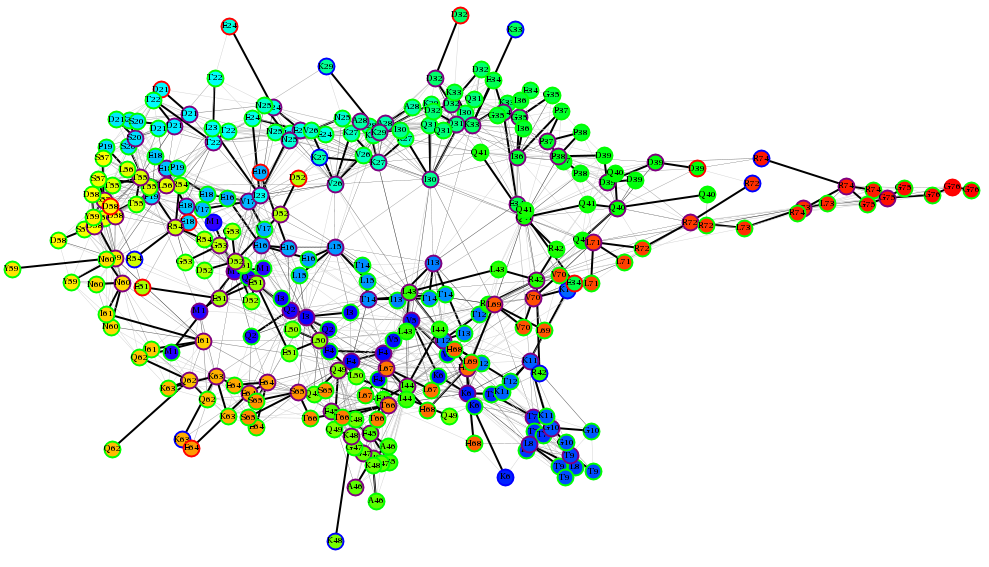 |
|
Benson NC, Daggett V (2012) A chemical group graph representation for efficient high-throughput analysis of atomistic protein simulations. J. Bioinformatics Compu. Biol. 0(2): 1250008-24. Benson NC, Daggett V (2012) A comparison of methods for the analysis of molecular dynamics simulations. J. Phys. Chem. B 116(29): 8722-31.
|
||
 |
Protein Flexibility Using principal component analysis, we can determine the a primary direction and magnitude of an atom's flexibility. By mining the entire Dynameomics database, we have discovered several interesting things about how proteins move and the relation of those movements to protein chemistry. |
|
Benson NC, Daggett V (2008) Dynameomics: large-scale assessment of native protein flexibility. Protein Sci. 17(12): 2038-50. |
||
|
|
||
Suite 328C
3401 Walnut Street
Philadelphia, PA 19104
Phone: 215-898-7300
Fax: 215-898-7301
email: nbe@sas.upenn.edu
