Social Influences on reproductive success in brown-headed cowbirds (Molothrus ater)
![]()
![]()
![]()
![]()
![]()
![]()
![]()
I have begun investigating ways in which the social environment can influence the development of animals' mating behavior. Historically, in investigations of development or learning, little is known about how animals actually acquire information from others in complex social settings. Thus, a primary goal of following studies was to determine if experiments, as opposed to descriptive studies, could be carried out on animals living in groups, continuously exposed to complex auditory and visual stimulation. A major concern was whether this multivariate approach to studying learning and development could be undertaken without sacrificing scientific rigor.

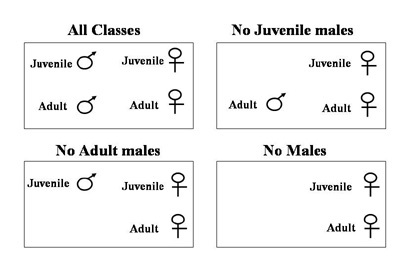
Using the outdoor aviary facilities at Indiana University and working in collaboration with Meredith West and Andrew King, we housed cowbirds in large social groups. To test the influence of cowbirds' social ecology on development, we used naturally occurring variation in social experience (the age class of individuals) to manipulate the social composition of these groups. In the first study, females were housed in one of four conditions: (1) with juvenile males, (2) with adult males, (3) with both juvenile and adult males, or (4) with no males.
During this experiment, I developed computerized system using voice recognition to record all data by speech and transcribe it to a database automatically. This allowed us to take extensive and detailed measures of birds' interactions daily throughout the year.
Developmental trajectories of all classes of cowbirds changed with social context.
Numerous differences in the birds' behaviors led to the emergence of dramatically different cultures in the four conditions as measured using auditory analysis, spatial assortment, and social interactions. The most pronounced effect was of adult male stimulation on juvenile male development. Juvenile males raised without adult males developed atypical social behavior.
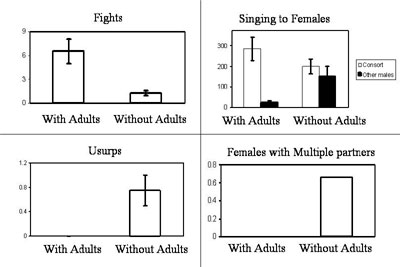
During the breeding season these males engaged in little intrasexual competition; they did not act aggressively toward one another nor did they guard females and they copulated with several different partners. These behavior patterns differed dramatically from juvenile males that had been raised with adult males. These juveniles behaved in a manner more typical of cowbirds in the Midwest; they were aggressive and socially monogamous. Furthermore, song analysis revealed that the songs of juveniles housed without adults were structurally different than that of juveniles housed with adult males, and were more effective in eliciting copulatory responses from females. Adult males appear to facilitate the development of competitive skills, but inhibit the learning or expression of vocal skills. These effects have now been replicated, found to be present very early in development, and persistent into adulthood. Furthermore, these patterns of behavior (both typical and atypical) can be culturally transmitted to a new generation of juvenile males.
We have recently studied the influence of females on the development of male mating behaviors. We had found previously that female affiliation with females and with males was influenced by male competition. Here we found that female behavior could control the development of male competition. Females that approached and affiliated with males more often stimulated male competition, which in turn stimulated female egg production. Female social proximity to juvenile males affected males' intrasexual competition, song potency, and courtship competency, copulation success and reproductive success.
Summary: Social factors were of fundamental importance to the expression of major aspects of cowbird behavioral repertoires. I have documented social influences on: copulation success, egg production, mating system, aggression, mate choice, mate preferences, song development, song use, and reproductive success. This paradigm, outlined in the above studies, provides a new means to study social influences on development, cognition, perception, attention, and learning as well as cultural evolution.
To read in more detail about this series of experiments, please visit the publications page of this site.
More information about the laboratory preparation:
I want to make clear that my work is a laboratory preparation and thus all the concerns of studying animals in captivity apply. The aviaries are indeed artificial; they are captive conditions and certainly can not be considered analogous to the wild. The housing conditions constrain subjects' range, they force individuals into the groups I create, birds are not exposed to predation, birds have access to food, water and shelter at all times. All of these factors and more could influence subjects' behaviour. I have moved to these types of conditions for my lab work instead of the more controlled lab settings typical of bird song experimental work because I have been influenced by the work of birdsong field biologists. Results of field studies often reveal that when birds are given the opportunity to have more degrees of freedom in their social behaviour, new patterns of learning emerge that can be very different from patterns seen in confined, socially isolated conditions. Even though my housing conditions do still impose constraints of the lab on my subjects, I believe that housing birds in groups provides a much more effective means to investigate their behavioral development.
When I began this work, I had concerns about studying cowbirds in confined conditions. I will discuss four issues that I considered as critically important when deciding to go forward with this preparation.
1.Using field data to guide experimental design. The variation that I study across social conditions occurs naturally across the cowbirds' range. I strive to create groups differing in social composition to mirror social compositions that have been reported in the wild. For example, in some western populations, adult birds move off their breeding areas quickly in search of food. In such populations, juvenile cowbirds may never interact with adults in their first year. In other locations, juveniles join flocks while adults are still in the final days of breeding and remain with the adults for their lifetimes. This variation served as the impetus for the White et al (2002c) study. In another example, in newly settled areas, cowbirds experience more social fluctuation with more individuals entering and leaving groups, whereas in other older populations there is more social stability.
It becomes clear when investigating field reports of cowbird social structure is that there is no longer a species-typical social composition. Cowbirds are non-territorial and range across the North American continent. Cowbirds seem to be able to exploit any type of habitat effectively and have moved into new areas as a result of human-induced habitat changes. Flocks composed of all females, all juveniles, and all classes have been reported in the field. There are also reports of variation across populations in: group density, breeding area climate, sex ratio, timing of the breeding season, and migration.
In addition to the variation in field reports of social ecology, there is also variation in field reports of cowbird behavior. This variation is similar to effects produced in my experiments. For example, reports of cowbird mating systems have ranged from promiscuity with no pair bonds, to female-defense monogamy. We generated this type of variability in White et al. (2002c). In addition, there are reports of regional differences in song dialects and whistles. Freeberg's work (1996, 1998), conducted in similar aviaries revealed that song dialects could be learned socially and could cause assortative mating in aviaries.
This variation is exactly what drew my interest to cowbirds. Cowbirds have moved into new ecological conditions rapidly and as such are often exposed to new social conditions. How is it that a species that seems to require social experience to develop normally can develop effectively in all sorts of different social environments? It is possible that a plastic developmental system could be the source of their ability to adapt so quickly to different environments.
2.The benefits of experimental control . Because of the artificial conditions, my studies cannot provide direct information about patterns of cowbird behavior in the wild. However, because I can influence social dynamics in a way that cannot be controlled in the field, I am able to have a window on behaviour that cannot be studied in the field. It is my hope that these lab findings will lead field investigations (see 3 below). I always construct the composition of the conditions to allow for comparisons across groups so that differences across conditions may reveal the variables that are important to cowbird behaviour. For example, I cannot and do not ever attempt to suggest that the number of eggs females lay in the aviaries is representative of the number of eggs females would lay in the wild because of the variables that differ in captivity (such as nutrition, calcium supplements, nest availability, etc). That being said, looking at individuals across years and across conditions reveals that certain characteristics of the social environment (male countersinging for example) play a role in influencing female decision processes for trading-off current and future reproductive success. This example also highlights the power of the aviary preparation, as effects like this could not be studied in the wild. In the wild it would be impossible (a) to find every egg females laid within or across years, and (b) to be able to compare patterns of egg laying across different types of social groups that do not differ in some other ecological variable.
3.Predictions for field study. In point one above, I discussed how using information from field reports is important in generating theory and experiments in the lab. It has, however, been extremely difficult to examine whether the variables that are revealing themselves as important in the lab are actually important in the field. This is a result of the difficulty of studying some of these early developmental experiences in a non-territorial species in the field. My work does provide a number of testable predictions for cowbirds in the wild. For example, results from White et al. (2002c) suggest that juvenile males living in flocks without adult males may experience less aggression in response to their directed song overtures. In the aviaries, one consequence of this was that juveniles without adult male exposure developed songs with more notes and more note clusters than did the juvenile males who were housed with adult males. Freeberg has studied the songs of two subspecies of brown-headed cowbirds, Molothrus ater artemisiae, and M. a. ater. Artemisiae males from his study population were more likely to be found in large all- juvenile flocks, whereas the ater juveniles were more likely to be found in mixed age-class flocks. Song analysis revealed that artemisiae males had significantly more note clusters and more notes within clusters in their songs than did the ater birds (Freeberg, 2001). This study reveals that it may be possible to study the social effects in which I am interested using aviary and field studies in an integrative manner.
4.Some benchmarks in aviary behaviour. Even though the above points argue for the use of cowbirds in my experiments, they do not speak to the issue of whether there may be problems with social housing of birds in these conditions. It is possible that confinement at such densities might produce abnormalities in behaviour such that results would not provide valid information about sexual selection or social learning. Here are some measures of behaviour in the lab that suggest I am not generating abnormal behaviour patterns.
Density . In Indiana , we studied several flocks in a very large aviary system (comprised of four smaller aviaries positioned end-to-end; spanning the length of a football field). While this is still smaller than their natural range, birds can disperse such that they would not be highly clustered. We have housed flocks in the large aviary complex four times now (including Smith, et al. 2000, for example), and each time we have done so, we have seen that birds did not disperse but instead clustered together in one quadrant of the large aviary in much higher densities than we had expected. The flock did move around in the large enclosure, and sub flocks formed, but bird always remain very closely clustered. This is at least suggestive that when they have more choice, they choose to remain in a group. Currently I am conducting an experiment designed to investigate how individuals housed in dynamic flocks (many individuals moving in and out throughout the year) compare to individuals housed in static flocks.
Wild caught birds . I am able to catch birds from the wild up to one month prior to the breeding season, place them in aviaries, and have them reliably sing, compete, court, mate, and reproduce. One of the main problems with captive breeding programs in other species is that the animals do not have the natural stimuli available to elicit reproduction. The aviaries provide males and females with the stimuli needed to reproduce. Wild caught adult males come into aviaries exhibiting the same degree of variation in social behaviour as the males raised in different social conditions in my experiments. In addition, females caught up to two months prior to the breeding season will respond in playback experiments and their patterns of responding with copulation solicitation displays to male songs are similar to the responses of females who have been in aviaries for their lifetimes. Also, in playback studies, responses by females to songs of wild caught males do not differ significantly from responses to songs of aviary-raised males.
False nests and female laying behaviour. Another abnormal aspect of the aviaries is that there are no host species available. I do make sure the false nests are as realistic as possible, using mock eggs in the nests (one is placed into a nest each day and new nests are added and old ones removed throughout the breeding season). I have now run a series of tests evaluating whether females are sensitive to the characteristics of these nests and have found that females evaluate egg size, shape, color and number (White et al, in press). Furthermore, they attend to nests and lay in patterns similar to those reported in the field. In the past we have experimented with hosts (juncos) in aviaries in the past, we never found any differences between patterns of laying when hosts were present and when absent. Cowbirds and juncos rarely ever interacted in the aviaries.
Finally, I have as little contact with the birds as possible. Birds are handled only rarely, and they are given ad lib food so that they do not associate humans with feeding schedules. I do not conduct steroid manipulations on the birds.
In sum, I strive to study variables that have been shown in the wild to be important to cowbird behaviour across their range. Cowbirds are an excellent species for studying social behaviour because there is such dramatic variability in social behaviour and social structure in the wild. While the data from the field certainly are indicative that social ecology affects learning, development and reproductive success, it is difficult to make such a causal statement without the experimental control provided by the lab environment.